
Nature: MERFISH绘制小鼠初级运动皮层空间细胞图谱
小鼠初级运动皮层(Mouse primary motor cortex,MOp)是额叶的一部分,主要负责计划、控制、运动执行,尤其是任何延迟反应有关的动作。在垂直方向上,皮层可以分为L1-L6六层,每层的形态和功能都有区别。大脑皮层的谷氨酸能神经元按其投射特性分为脑内(IT)神经元、大脑下投射神经元(或锥体束神经元)和皮质丘脑投射(CT)神经元;γ-氨基丁酸(Gaba)能神经元可以根据发育起源于内侧神经节隆起和尾神经节隆起进行分类,并可以通过小白蛋白(Pvalb)、生长抑素(Sst)、血管活性肠肽(Vip)和Lamp5等标记基因进一步分类。单细胞转录组学研究揭示了大脑细胞的高度多样性,并报道了在单个皮层区域中存在数十到上百种细胞类型。然而,想要了解不同类型细胞对大脑功能的作用仍然缺乏一个高分辨率、包含细胞空间组织和连接信息的细胞图谱。庄小威团队使用MERFISH(multiplexed error-robust fluorescence in situ hybridization,多路抗错荧光原位杂交)来识别不同的细胞类型,并绘制它们在小鼠初级运动皮层(MOp)中的空间图谱。通过整合MERFISH和神经逆行标记,进一步揭示了不同类型细胞的投射模式的复杂性。
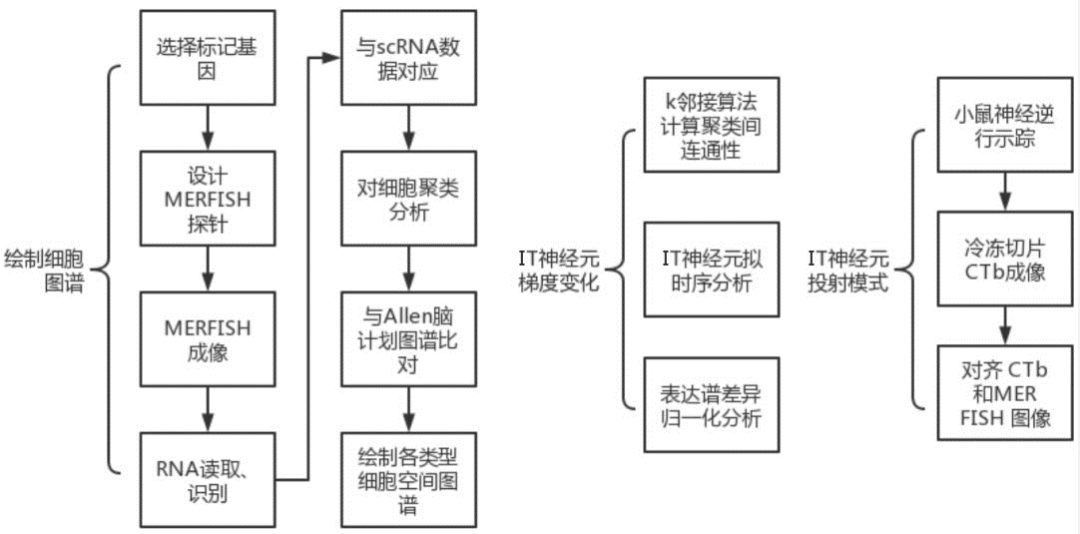
1.1 MERFISH成像及细胞聚类分析
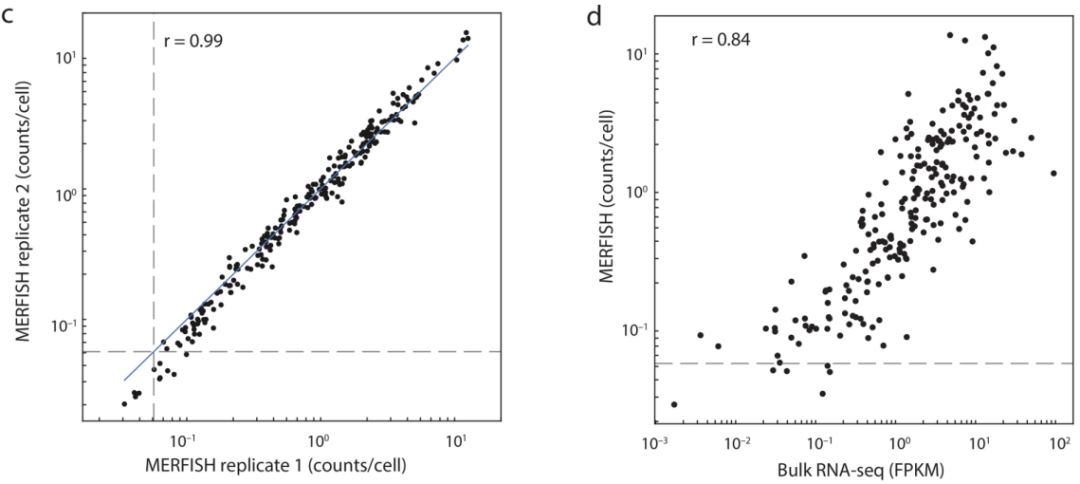
作者选择了258个标记基因进行MERFISH成像,其中包括已知的大脑皮层中主要神经元和非神经元细胞类型的典型标记基因,以及由单细胞和单核RNA测序联合鉴定的神经元簇中差异基因表达以及互作分析选定的标记基因。MERFISH成像后将mRNA分配到单个细胞中,258个基因中有4个染色“不合格”,在随后的分析中排除。从MERFISH中得到的单个细胞平均基因拷贝数在其他小鼠中可以重复,并且与混合RNA-seq测定的基因表达水平具有高度的相关性(图1)。之后,通过无监督的聚类分析的MERFISH衍生的单细胞表达谱鉴定了39个兴奋性神经元簇,42个抑制性神经元簇和14个非神经元簇(图2)。
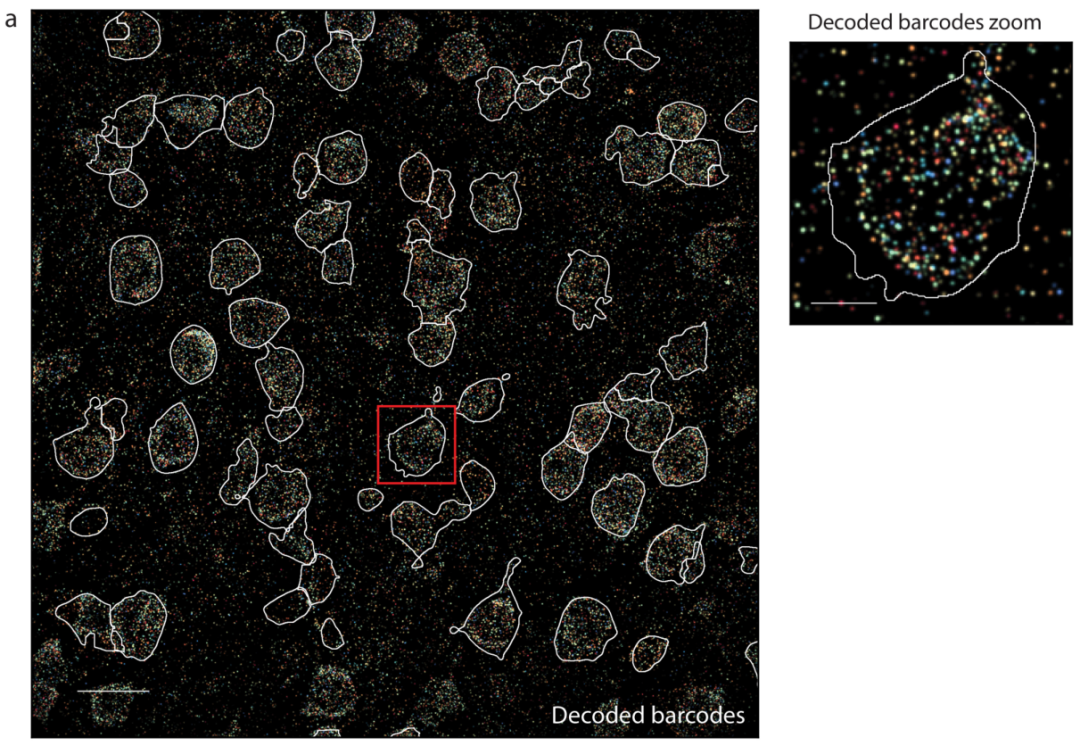
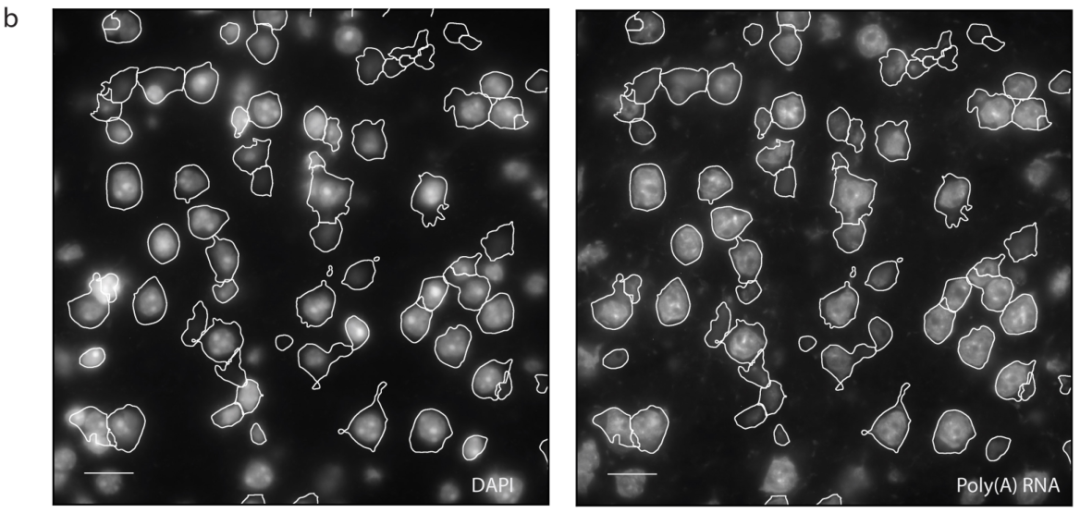
a,单个视场的解码MERFISH图像,白线为识别的细胞。比例尺,20 μm(左)和 5 μm(右)。b,DAPI(左)和poly(A)RNA(右)图像。
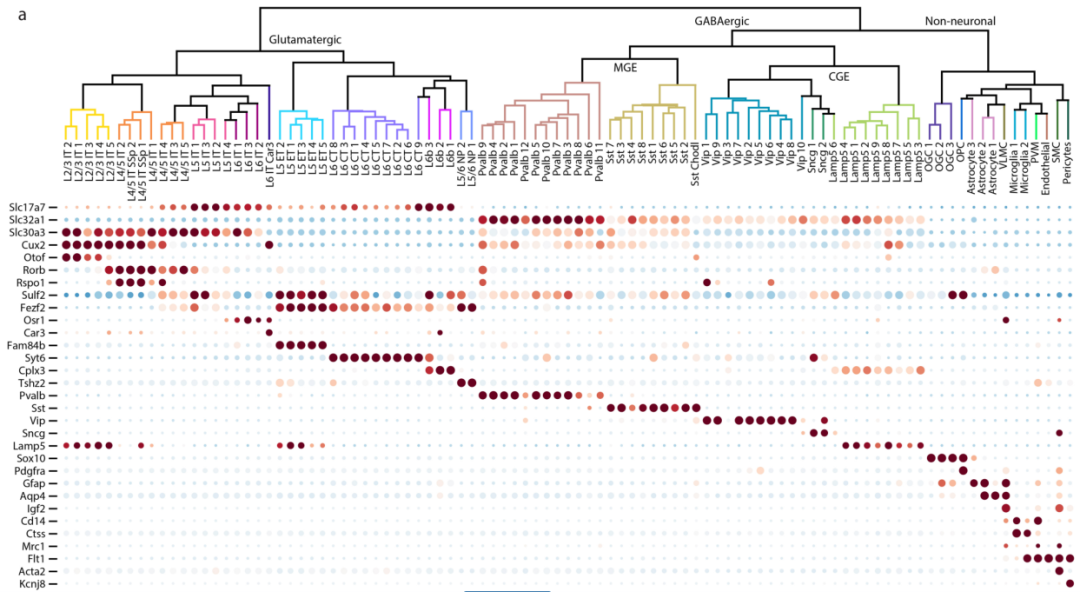
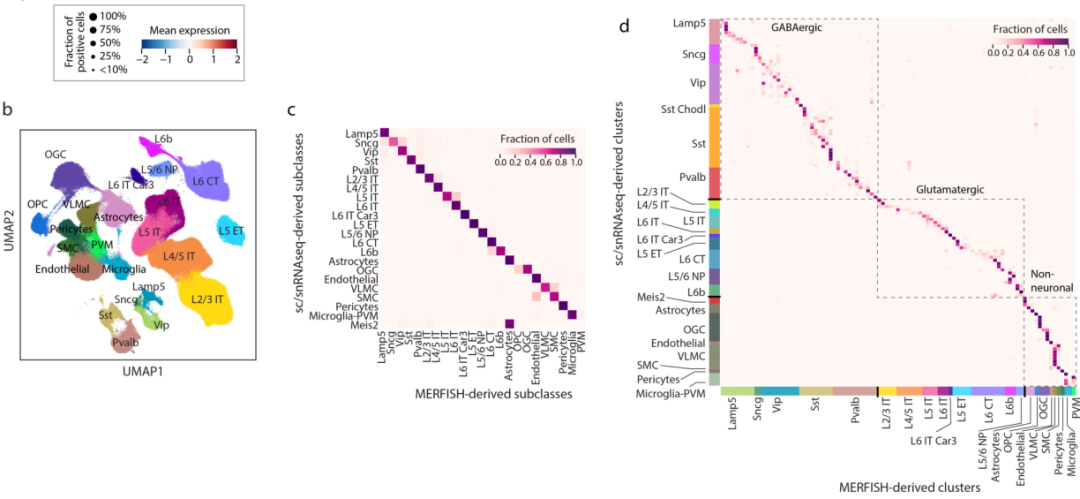
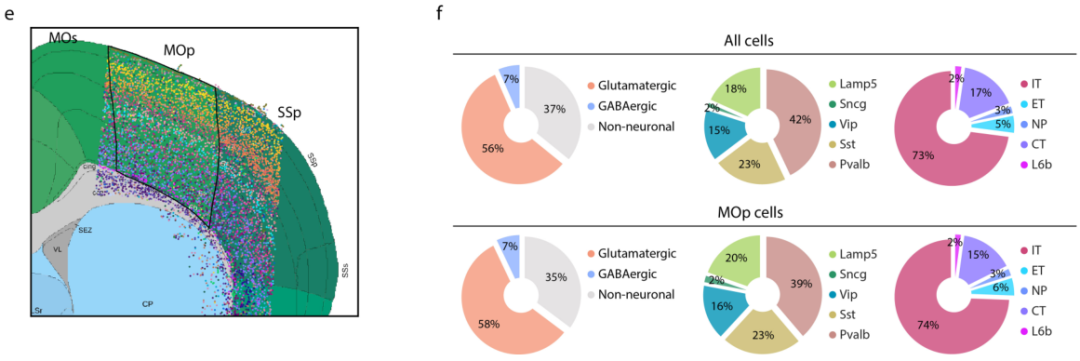
图2基于MERFISH的细胞类型分析,基于共同坐标框架的MERFISH图像配准和细胞组成
a,树状图显示了MERFISH鉴定的39个谷氨酸能簇、42个gaba能簇和14个非神经元簇之间的层次关系。b,用UMAP细胞类型进行分类。c&d,MERFISH 确定的细胞亚类与scRNA-seq 和snRNA-seq数据集确定的细胞亚类之间的对应关系。e,MERFISH对不同细胞簇染色示意。f,整个成像区域以及初级运动皮质(MOp)中三个主要细胞类别。
1.2细胞分布的空间差异
MERFISH 图像直观地显示了在 MOp 及其邻近区域细胞的转录差异(图3 a)。谷氨酸能细胞簇,特别是IT亚类,在全部分层都有分布(图3b)。与几乎跨越整个皮层的IT细胞不同,ET、NP、CT和L6b细胞只分布在较深的分层(图3 b,c)。个别的谷氨酸能细胞簇沿着皮层方向呈现出区分明显的、部分重叠的分布,而且这些细胞簇许多呈现分布宽度小于个别皮层的厚度。Gaba能神经元也表现出高水平的空间多样性,Lamp5、Sncg和Vip亚类在表层更多,而Sst和Pvalb亚类在深层更多(图3b,d) 。值得注意的是,在亚群水平,大多数Gaba能亚群优先分布在一个或两个皮质层(图3d)。作者还绘制了非神经元细胞的空间组织(图3b,e)。在三个星形胶质细胞群中,星形胶质细胞1分布于各层,星形胶质细胞2分布于L1和白质中,星形胶质细胞3几乎只分布于白质中。少突胶质细胞谱系分为少突胶质前体细胞和3个成熟少突胶质细胞群,成熟少突胶质细胞在白质中富集,少突胶质前体细胞均匀分布于各层。血管软脑膜细胞形成皮层的最外层细胞。其他类型的非神经细胞在皮质层和白质中的分布更为分散。
作者注意到不同细胞种群在空间上大量混合。为了量化每个细胞邻近区域的细胞组成的复杂性,确定了每个细胞邻近区域存在的不同细胞簇的数量,并观察到高水平的局部细胞异质性,细胞邻近区域的组成复杂性向深层递增(图3f)。
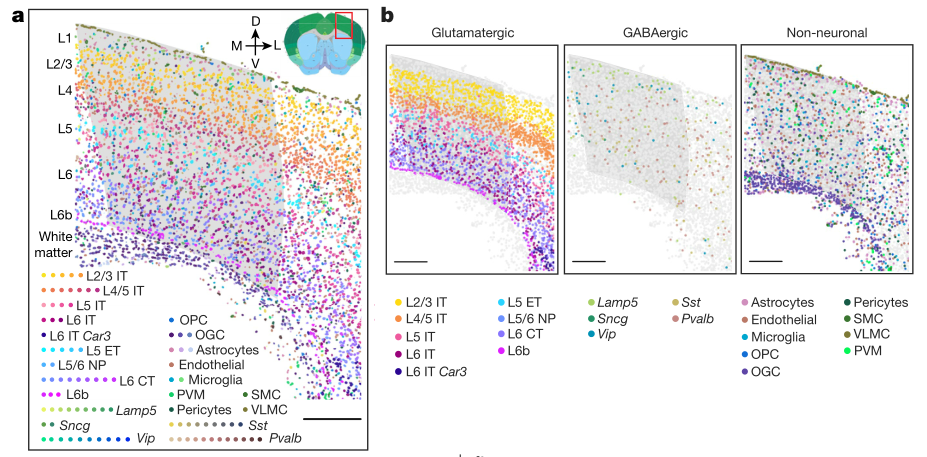
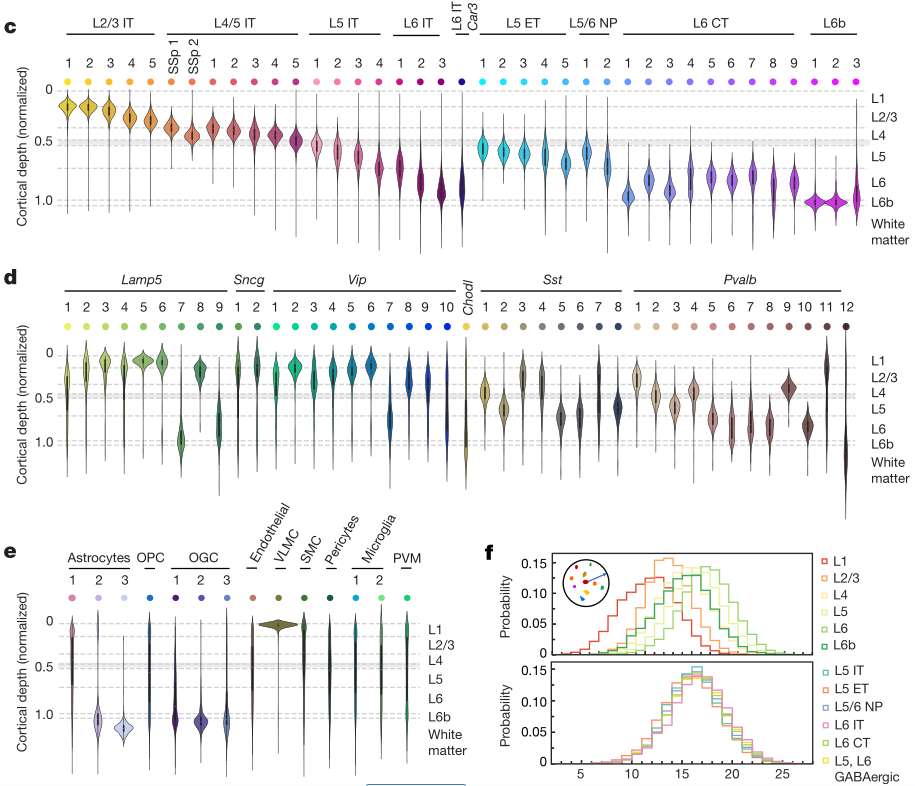
a,冠状切片中细胞群的空间图。基于Allen图谱确定的MOp区域是灰色的。b,相同切片的谷氨酸能(左)、GABA能(中)和非神经元(右)细胞亚类的空间分布图: 细胞呈圆形,指示细胞呈亚类染色,其他细胞呈灰色。比例尺,400μm。c-e,谷氨酸能神经元、非神经元细胞、整个成像区域的细胞簇在皮质深度的分布。f,L5和L6皮层(顶层)和不同亚类细胞邻域复杂度的概率分布。细胞的邻域复杂度定义为在给定细胞周围半径100μm范围内存在的不同细胞簇的数目。
综上所述,作者使用 MERFISH 生成了小鼠MOp及其邻近区域的分子定义和空间解析的细胞图谱。使用MERFISH对约30万个细胞进行分析,识别出95个神经元和非神经元的细胞群,并揭示了大多数转录不同的细胞群的不同空间分布。作者的研究结果显示了不同亚类神经元分布的层级限制,并揭示了先前未知的个别神经元簇的高分辨率空间映射。作者观察到层状组织不仅包括兴奋性神经元,而且也有抑制性神经元,许多抑制性神经元簇优先位于一个或两个皮层。此外,许多兴奋性神经元簇沿皮层深度方向呈狭窄分布,显示出个别皮层内较细小的层状结构。
作者注意到,尽管神经元倾向于在亚类水平上形成表达谱不同的离散细胞群,但是单个亚类中的细胞簇往往表现出更多的渐变,这证明了脑细胞的离散和连续异质性共存。特别是占MOp兴奋性神经元总数的70% 左右的IT神经元,在大脑皮层深处的分布形成一个连续的梯度。同时进行的scRNA-seq研究也观察到同位皮层 IT 神经元之间基因表达的持续变化。通过空间解析的单细胞分析,作者观察到 IT 神经元的基因表达随皮层深度的相关变化。
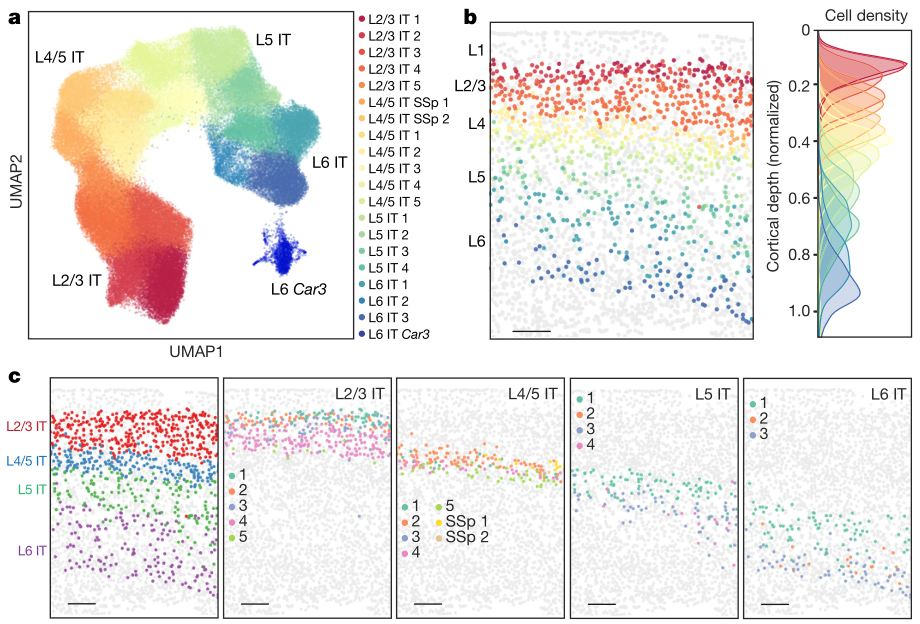
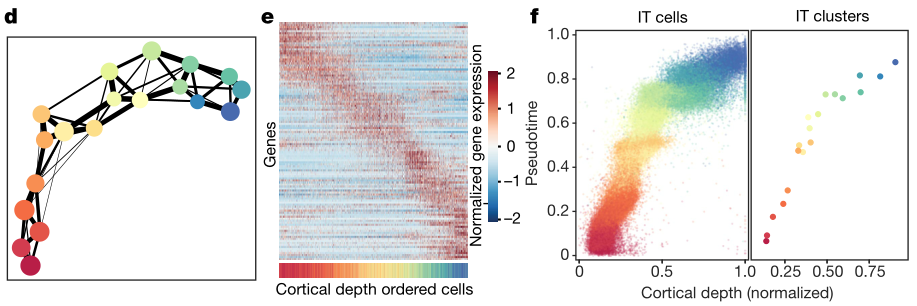
a,对整个成像区域的 IT 神经元UMAP分析。b,冠状切片上IT神经元的空间分布图。IT 神经元像 a中被它们的集群特性所影响,而其他所有的细胞都是灰色的(左)。比例尺,200μm。皮质深度分布的个别 IT 集群也显示(右)。c,在同一冠状切片上,每个亚类(左侧面板)以及 L2/3、 L4/5、L5和L6 IT亚类(4个右侧面板)中单个簇的空间分布图,比例尺,200μm。d,IT集群的 k- 近邻图中集群之间的连通度,每个集群用 a 中的彩色节点表示,节点之间的加权边表示连通度。e,所有 IT 神经元跨皮层深度差异表达基因的标准化表达。在这里,差异表达基因是指表达基本不同的皮层深度(方法)。个别的 IT 神经元按照皮层深度上升的顺序分类,基因按照表达最强的皮层深度分类。底部的颜色条显示了细胞的集群特性。f,散点图显示了被细胞集群染色的L2/3,L4/5,L5和L6的单个IT神经元(左)和单个 IT 集群(右)的拟时序与正常皮质深度的关系。
作者将霍乱毒素b亚单位(CTb)注射到三个皮层区域,即同侧 MOs 区、SSp 区和颞关联区(TEa) ,通过 CTb 成像识别MOp 中投射到这些靶区的神经元。通过结合MERFISH和神经逆向标记技术,作者进一步研究了个别的细胞类型与其投射靶点之间的关系。研究结果表明,MOp 神经元向其他皮层区域的投射形成了一个复杂的多对多网络: 每个细胞簇投射到多个靶区(与之前对视觉皮层投射的观察结果一致) ,每个靶区接收来自多个细胞簇的投射。作者还观察到一些相似的神经元簇具有不同的投影特性,这些神经元簇具有逐渐变化的表达谱和重叠的空间分布。作者测量了三个目标区域,而更多的目标区域可以用这种方法来测量,以构建一个更全面的投影图的细胞类型的MOp。作者设想 MERFISH 也可能与跨突触病毒示踪剂结合,生成一个高分辨率的细胞类型到细胞类型的连接图。
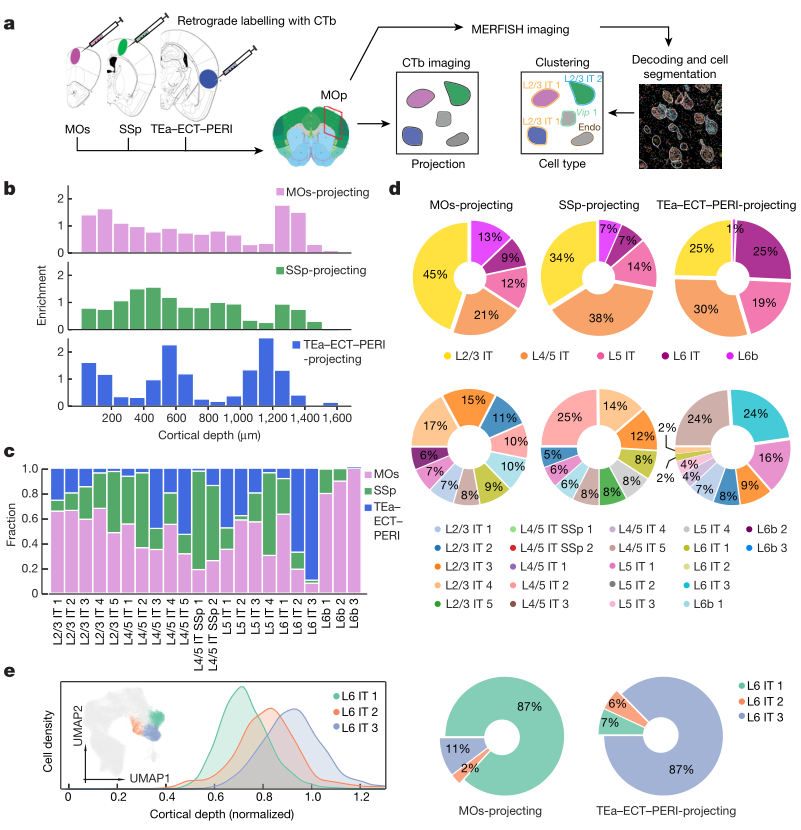
a,逆行标记和MERFISH整合的工作流程。b,在不同皮层深度富集 MOs-projecting、SSp-projecting 和 TEa-ECT-PERI-projecting 细胞。富集定义为相关 CTb 阳性细胞的分数除以同一bin中所有 IT 和 L6b 细胞的分数。c,在簇中所有 CTb 阳性单投射细胞中,每个细胞簇中 MOs 投射、SSP 投射和 TEa-ECT-PERI 投射细胞的分数。d,饼图显示属于每个细胞亚类(顶部)和簇(底部;仅前 10 个簇)的 MOs 投射(左)、SSp 投射(中)和 TEa-ECT-PERI投射(右)细胞的比例。e,分子和空间相似的 L6 IT 簇的投影特异性。显示了 L6 IT 1-3 簇和 UMAP(插图)的皮层深度分布,L6 IT1-3神经元如图例颜色,其他 IT 神经元以灰色显示(左)。饼图显示了属于三个簇中的每一个的 MOs 投射和 TEa-ECT-PERI 投射 L6 IT 神经元的相对比例(右)。
有趣的是,某些分子水平和空间水平相似的 IT 团簇呈现出不同的投射模式。例如,几乎所有CTb阳性的 L6 IT3神经元都投射到 TEa-ECT-PERI 上,但没有投射到MOs和SSp上,而CTb阳性的 L6 IT1神经元大部分投射到MOs上,只有极少数投射到 TEa-ECT-PERI (图5 c)。尽管三个 L6 IT 簇的基因表达谱相似,空间分布也基本重叠,然而MOs主要接受 L6 IT 1的输入,而TEa-ECT-PERI则主要接受L6 IT 3的输入(图5 e)。
本文使用单细胞转录组成像技术——多路抗错荧光原位杂交(MERFISH),生成一个小鼠初级运动皮质空间解析细胞图谱。作者在小鼠初级运动皮层及其邻近区域分析了大约30万个细胞,鉴定了95个神经元和非神经元细胞簇,揭示了一个复杂的空间图谱。研究发现不仅是兴奋性神经元细胞簇,而且大多数抑制性神经元细胞簇都采用层状分布。脑内(IT)神经元沿皮层深度轴形成了连续的梯度变化,其单个细胞的基因表达与其皮层深度相关。此外,作者将MERFISH与逆行标记相结合,探测小鼠初级运动皮质(MOp)神经元的投射目标,发现它们的皮层投射形成了一个复杂的网络,单个靶区接收来自多个神经元簇的输入,单个神经元也会投射到多个靶区。

Zhang M,Eichhorn SW, Zingg B, et al. Spatially resolved cell atlas of the mouse primarymotor cortex by MERFISH. Nature. 2021;598(7879):137-143.
doi:10.1038/s41586-021-03705-x
最新动态
-
11.15
【干货】手把手教你做酵母双杂交点对点验证
-
11.14
酵母双杂交点对点验证怎么做?及常见问题回答
-
11.13
酵母双杂交建库-试剂盒操作流程解析及常见问题解答
-
10.17
一文了解 | 酵母双杂交建库原理及流程
-
10.14
盘一盘 诺奖明星-micro RNA的前世今生
-
09.30
干货图文| 双荧光素酶报告基因结果解读
-
09.23
中药的现代诠释:外泌体如何革新传统医学?
-
09.10
【客户文献分享】SGLT2 抑制剂达格列净可改善高尿酸血症性肾病的肾脏纤维化
-
08.30
客户文章分享 | PTPRE抑制剂有望成为治疗HCC的新策略
-
08.29
知无不“研” | 一文透彻理解荧光素酶蛋白互补实验LCA
 X
X 






