
干货| 一文读懂并区分双荧光素酶实验(DLR)、双分子荧光互补(BiFC)、荧火素酶互补实验( LCA)
作为现代生物学研究的重要工具,荧光技术凭借其高灵敏度和非侵入性特点,在科研领域发挥着不可或缺的作用。然而,对于许多初学者来说,双荧光素酶实验(DLR)、双分子荧光互补(BiFC)以及荧光素酶互补实验(LCA)这三种荧光实验技术却常常因为名字相似而难以区分。这不仅可能导致在实验设计时的混淆和误解,还可能浪费大量的时间和资源在不必要的重复实验上。本文旨在深入剖析三种重要荧光实验技术,通过直观的对比和文献案例,让您轻松掌握它们的核心原理、差异与实战应用,从而提高研究效率和质量。
一、双荧光素酶实验(DLR)
►原理:在双荧光素酶实验中,将萤火虫荧光素酶作为实验报告基因,海肾荧光素酶作为对照报告基因。实验报告基因用于测试实验条件下基因的表达,而对照报告基因作为内对照。以荧光素为底物来检测萤火虫荧光素酶活性,荧光素酶可以催化荧光素,在荧光素氧化的过程中会发出生物荧光,然后通过化学发光仪测定。
①转录因子与启动子
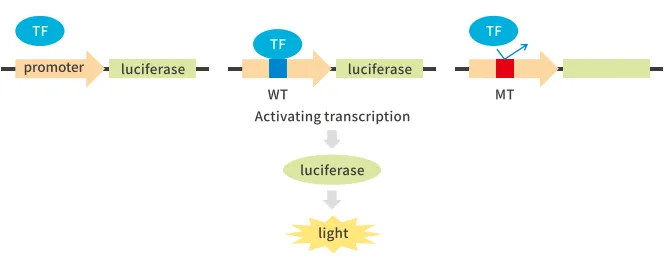
将启动子序列构建到萤火虫荧光素酶基因前,同时过表达转录因子。当转录因子与启动子上特异结合位点结合后激活荧光素酶基因转录,使萤火虫荧光素酶得以表达,最终荧光强度上升;当结合位点被突变后,转录因子无法与启动子结合,因此荧光值无明显变化。
②miRNA与靶基因
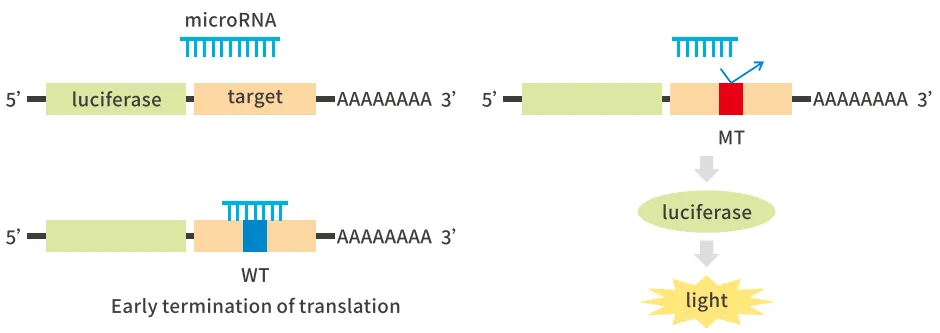
将靶基因序列构建到萤火虫荧光素酶基因3'区域,同时过表达microRNA。当microRNA与靶基因上特异结合位点结合后,将干扰荧光素酶mRNA的翻译或导致其迅速降解,使荧光强度降低;当结合位点被突变后,microRNA无法与靶基因结合,因此荧光值无明显变化。
►实验流程
1.根据组学分析结果确定miRNA与靶基因/转录因子与启动子相关信息,或者软件预测结合位点。
2.基因合成靶基因序列,将该片段构建到萤火虫报告基因载体,测序验证。
3.基因合成pre-miRNA或转录因子序列,将该片段构建到过表达载体,测序验证;或使用合成的mimics进行实验。
4.将报告基因质粒和转录因子或pre-miRNA过表达质粒共转染目标细胞或烟草。
5.裂解细胞用于荧光素酶检测。
6.加入酶作用底物,化学发光仪测定荧光素酶的活性。
7.数据计算及结果分析。
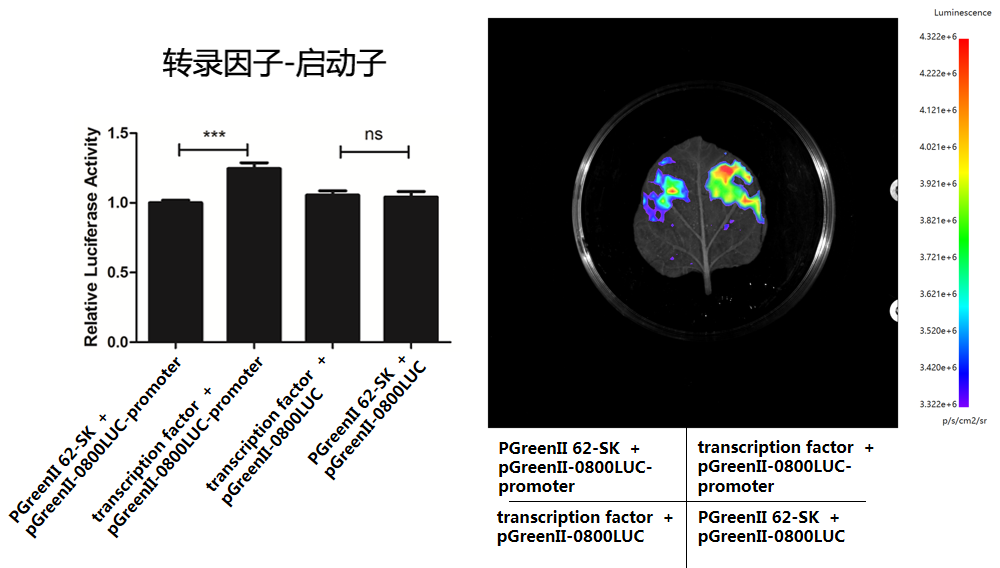
►结果展示
二、双分子荧光互补(BiFC)
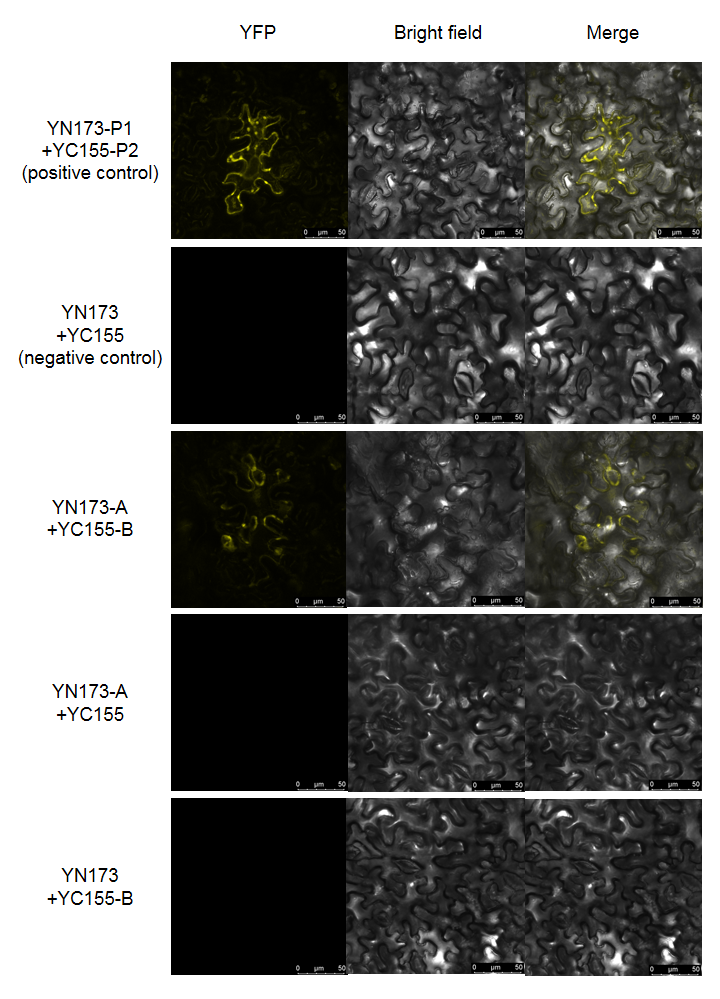
►原理:该技术将荧光蛋白分割成两个不具有荧光活性的片段(N端和C端),然后将这两个片段分别融合到两个目标蛋白上。当两个目标蛋白在细胞内相互作用时,荧光蛋白的两个片段在空间上靠近并重新组合成完整的荧光蛋白,从而发出荧光。
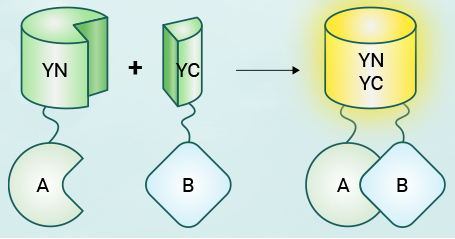
►实验流程
● 构建融合蛋白:将待测蛋白A和B分别与荧光蛋白N端和C端片段基因融合,构建相应的表达载体。
● 烟草转化:将含有融合蛋白A-N端和B-C端片段的载体同时转入烟草中,实现融合蛋白的共表达。
● 荧光观察:转染后一段时间(通常24-48小时),使用荧光显微镜或共聚焦显微镜观察细胞,若观察到绿色(或对应荧光蛋白颜色)荧光信号,说明蛋白质A和B发生了相互作用。
►结果展示
三、荧火素酶互补实验(LCA)
►原理:将萤火虫荧光素酶蛋白分成两个片段,分别连接两个待测目标蛋白,当两个目标蛋白相互作用时,荧光素酶的两个片段在空间上靠近并重新组合成完整的荧光素酶,从而催化荧光素产生荧光。
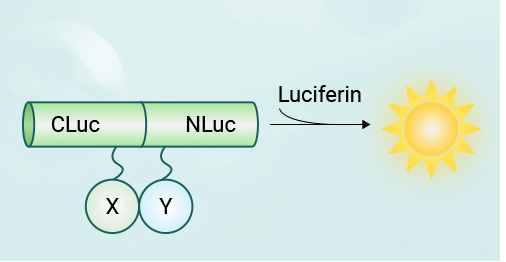
►实验流程
● 构建融合蛋白:选取待研究的两个蛋白质(A与B),将荧光素酶裂分为N端和C端非活性片段,分别与蛋白A和B融合,构建表达载体。
● 烟草转化:将载体转化到烟草叶片中,确保蛋白表达。
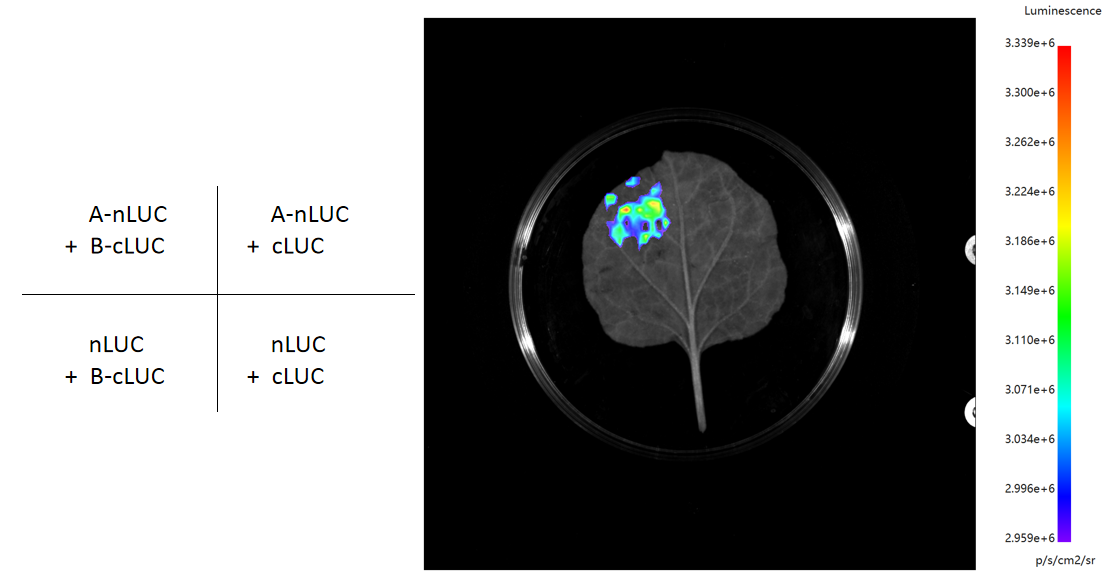
● 荧光检测:添加荧光素酶底物,若蛋白A与B相互作用,重组的荧光素酶将催化发光,通过成像系统捕捉荧光信号。
● 图像分析与验证:分析荧光信号,设立对照实验验证结果,排除非特异性信号,确定蛋白质相互作用的空间与时间模式。
►结果展示
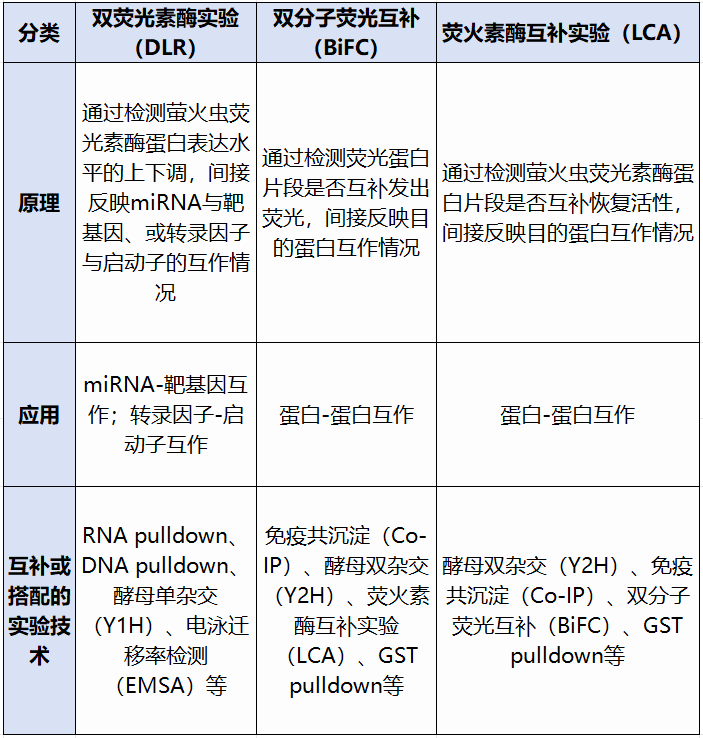
在深入探讨了双荧光素酶实验(DLR)、双分子荧光互补(BiFC)技术以及荧火素酶互补实验(LCA)的原理和实验流程后,相信大家已经对这三种技术有了较为清晰的认识。为了进一步帮助读者朋友们理解这些技术的特点和具体应用,小编将通过一张简明的表格来对比它们之间的原理、应用以及互补或搭配的实验技术。
需要注意的是,这些搭配的实验并不是绝对的,而是根据实验条件和目标来选择的。在实际研究中,研究者需要根据具体情况选择合适的实验方法。
理论告一段落,接下来,小编将展现双荧光素酶实验、BiFC及LCA在实际研究中的应用案例,让大家更直观地理解这些技术如何在科学研究中发挥作用。
案例一:转录因子VaNAC72调控葡萄VaCP17基因表达以提高耐寒性

植物需要自我调整以应对寒冷、干旱和盐碱等胁迫的发生。寒冷胁迫严重威胁植物的正常生长、发育和产量,因此阐明植物对寒冷反应的内在机制和挖掘冷胁迫调节级联将有助于抗寒性的分子育种。
本研究通过VaCP1在冷敏葡萄“无核白”中的稳定过表达和VaCP17在“双优”叶中的瞬时沉默,研究了VaCP17的耐寒功能。随后,作者通过酵母单杂交(Y1H)、双荧光素酶报告基因检测(DLR)和VaNAC72在“双优”叶片中的瞬时过表达分析了VaNAC72与VaCP17的调控关系。此外,作者还通过酵母双杂交(Y2H)、双分子荧光互补(BiFC)和荧光素酶互补(LCA)等方法鉴定了与VaNAC72相互作用的蛋白,并通过DLR进一步分析了VaNAC72与相互作用蛋白之间的共调控关系,旨在揭示VaCP17介导的耐寒性的分子机制。
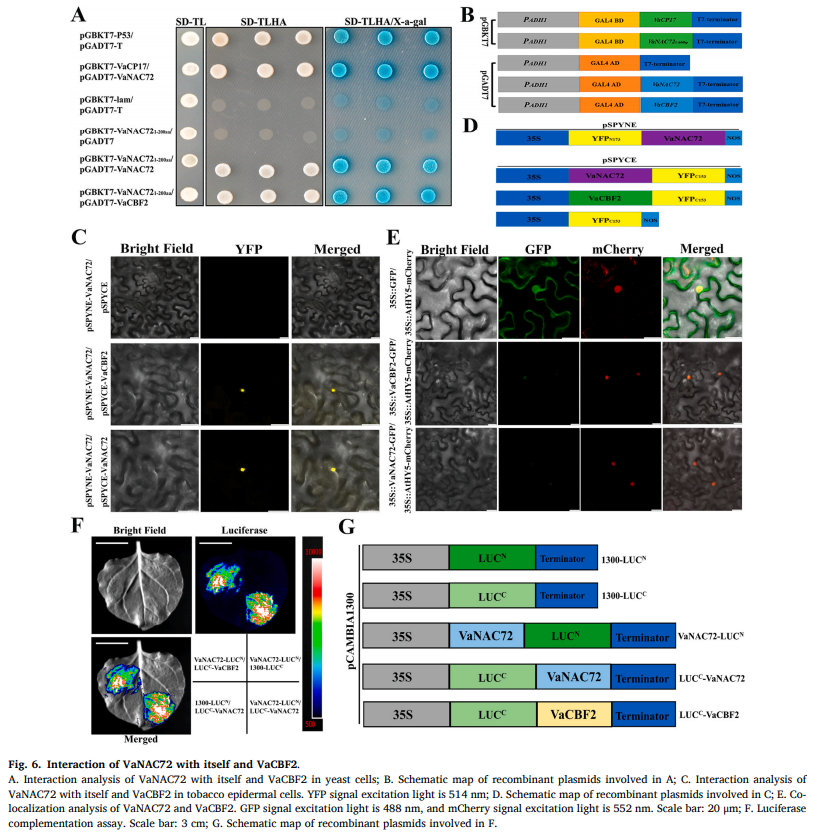
图源:https://doi.org/10.1016/j.plaphy.2023.
案例二:MdERF114正向调控苹果根系抵御腐皮镰孢菌侵染的分子机制

研究发现,MdERF114在苹果根系中受腐皮镰孢菌诱导高度表达。过表达MdERF114提高了苹果根系对腐皮镰孢菌的耐受性,而MdERF114干扰根系则表现出对腐皮镰孢菌敏感。作者通过酵母单杂交、双荧光素酶和EMSA等技术发现转录因子MdERF114通过结合MdPRX63启动子并调控其转录促进苹果根系中木质素的沉积,从而提高对腐皮镰孢菌的抗性。研究还发现MdWRKY75受腐皮镰孢菌诱导上调,并结合MdERF114启动子的W-box元件调控MdERF114的表达,从而获得对腐皮镰孢菌的抗性。
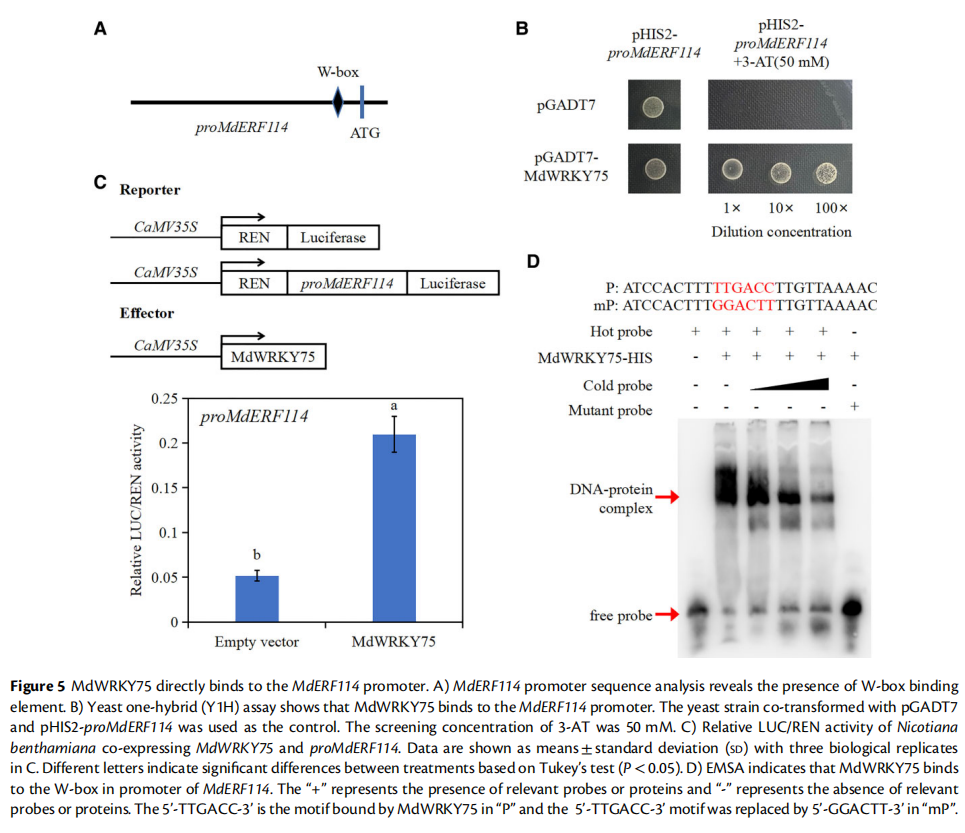
图源:https://doi.org/10.1093/plphys/kiad057
此外,作者通过酵母双杂交、荧光素酶互补实验和双分子荧光互补等实验证实MdMYB8与MdERF114存在相互作用,参与MdERF114介导的对腐皮镰孢菌的防御网络。总的来说,该研究为苹果根系响应腐皮镰孢菌侵染的调控机制提供了新的见解,为培育抗性砧木和防治苹果重茬连作障碍奠定了理论基础。
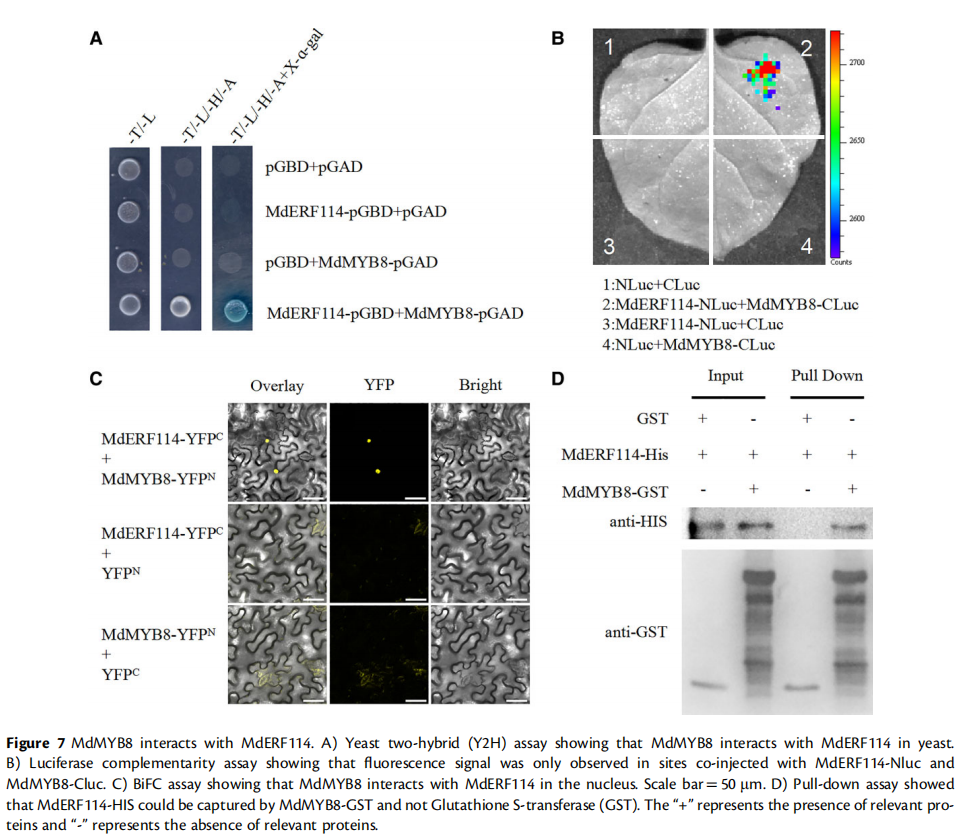
图源:https://doi.org/10.1093/plphys/kiad057
案例三:茉莉酸反应性转录因子NnWRKY70a和NnWRKY70b正调控荷花中苄基异喹啉生物碱的生物合成

在该项研究中,作者通过:(1)系统发育分析表明,NnWRKY70a 和 NnWRKY70b 为典型的可参与植物次生代谢调节的III 组 WRKY蛋白;(2)双荧光素酶和酵母单杂交实验表明,NnWRKY70a 和 NnWRKY70b 可以特异性结合BIA 途径基因启动子的 W-box顺式元件,从而正向调节荷花 BIA 生物合成,NnWRKY70b 的 BIA 生物合成激活比 NnWRKY70a 更强;(3)酵母双杂交和 BiFC 分析进一步揭示NnWRKY70b通过与在调节荷花 BIA 生物合成中的关键蛋白之间的相互作用,调节荷花 BIA 生物合成。以上的研究结果证明了 NnWRKY70 TFs 在激活荷花中 BIA 的生物合成中的积极调节作用,并为通过基于 TF 的基因工程提高 BIA 产量提供了可行的策略。
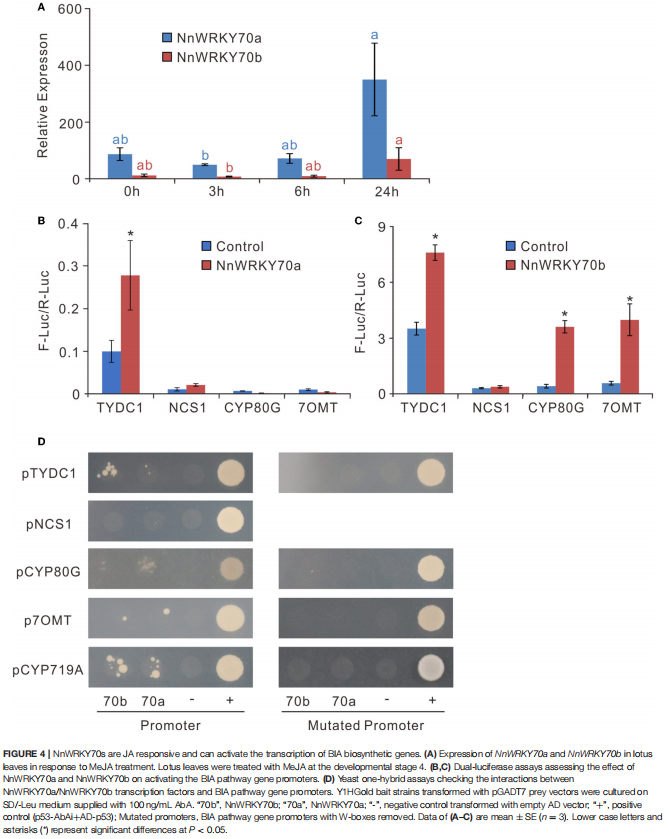
图源:doi: 10.3389/fpls.2022.862915
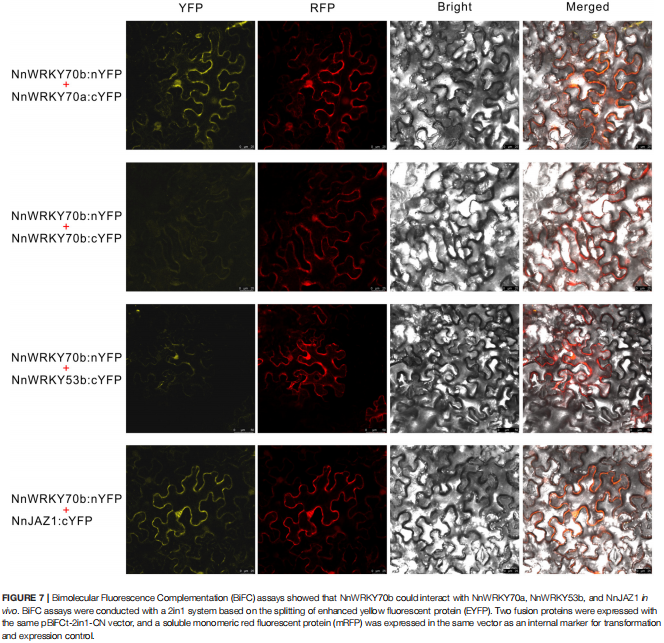
图源:doi: 10.3389/fpls.2022.862915
双荧光素酶实验(DLR)、双分子荧光互补(BiFC)和荧光素酶互补实验(LCA)虽名称相近,但各有千秋,理解这些技术的基本原理与应用场景,对于科研工作者而言至关重要。
最新动态
-
03.11
同为抗体,为什么一抗看“抗原种类”,二抗却看“一抗种属”?
-
03.11
J Nanobiotechnology(IF=12.6):蔓越莓“纳米邮差”递送麦角甾醇,重启早衰卵巢功能!
-
02.27
没有抗体?不是模式植物?DAP-seq照样搞定你的TF靶基因!
-
02.27
高分研究的共同选择:三篇权威文献背后的ChIP实验工具解析
-
02.27
表观遗传学进入“多维验证”时代,这些疾病机制再也藏不住了!
-
02.27
“药食同源”顶刊思路(IF=10.5)!来源鉴定+活性追踪+体内验证,全面解析无花果来源外泌体抑制乳腺癌骨转移的双重机制!
-
02.27
千年古方,纳米新生:科学家破解半夏如何“指挥”免疫细胞对抗肺癌!
-
01.29
技术跨界联用:表观遗传学进入“多维验证”时代,这些疾病机制再也藏不住了!文末有惊喜活动!
-
01.29
机制研究的金标准:金开瑞RIP/RNA pull-down试剂盒获多篇顶刊研究认可!
-
01.29
想测结合常数,但样本太珍贵?来试试MST技术
 X
X 






