
转录组、蛋白质组学联合案例分析
信息来源:金开瑞 作者:genecreate 发布时间:2018-08-29 16:06:38
标题:Transcriptomic, Proteomic, and Bioelectrochemical Characterization of an Exoelectrogen Geobacter soli Grown With Different Electron Acceptors
用不同电子受体培养的产电菌群Geobacter soli Grown的转录组学、蛋白质组学和生物电化学表征
期刊:Front Microbiol
影响因子:4.019
研究背景
Geobacter是一种发电菌属,其细胞外电子转移(EET)能力在生物地球化学和生物能源过程中发挥重要作用,作者选择Geobacter soli GSS01菌种,利用转录组学、蛋白质组学技术希望揭示EET在不同Geobacter菌种之间的差异。
研究结果
1. G.soli的细胞外电子转移(EET)能力
作者培养了G.soli和G.sulfurreducens PCA两种菌株,发现这两种菌株都能够还原四种常见的不溶性Fe(III)氧化物。随着培养时间的延长,GSS01菌株还原生成的Fe(II)产物量比PCA菌株多,这些结果表明GSS01菌株可能比PCA菌株更快地还原Fe(III)氧化物。在电流密度和电流强度方面,GSS01菌株也高于PCA菌株。
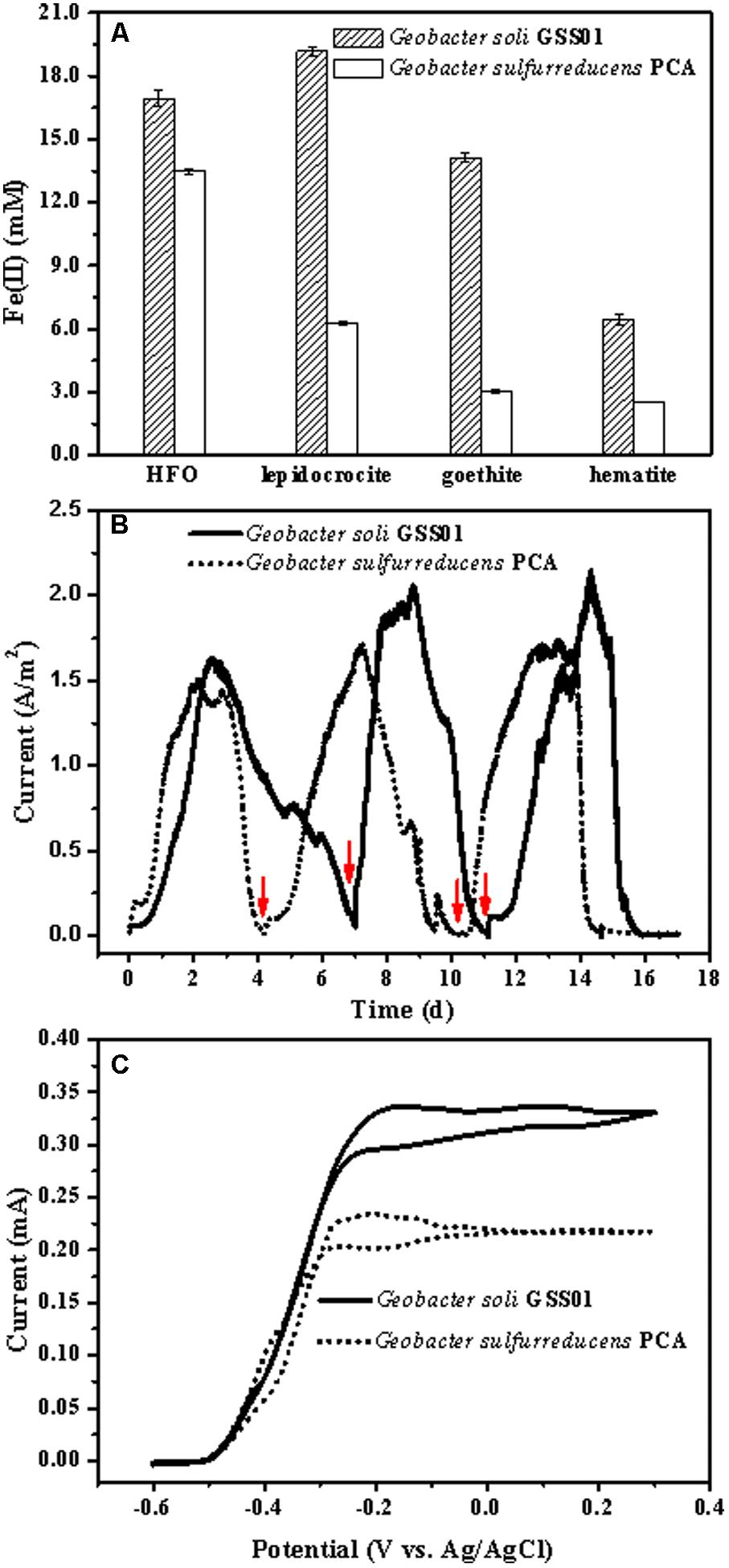
图1 G.soli GSS01和G.sulfurreducens PCA 的EET能力比较
2. G.soli生物膜的生物电化学特征
为了准确识别出负责发电的生物催化活性位点,作者使用non-turnover CV法和turnover DPV法分析菌株生物膜,发现两种生物膜均显示出两个主要的氧化还原状态(E2、E3)和一种次要氧化还原状态(E1)。而且G.soli生物膜中也存在一个独特的次要氧化还原状态,其形式电位为-0.20 V(E4)。除此之外,EIS分析显示,G.solid 生物膜具有比G.sulfurreducens更低的电荷转移电阻(Rct)。
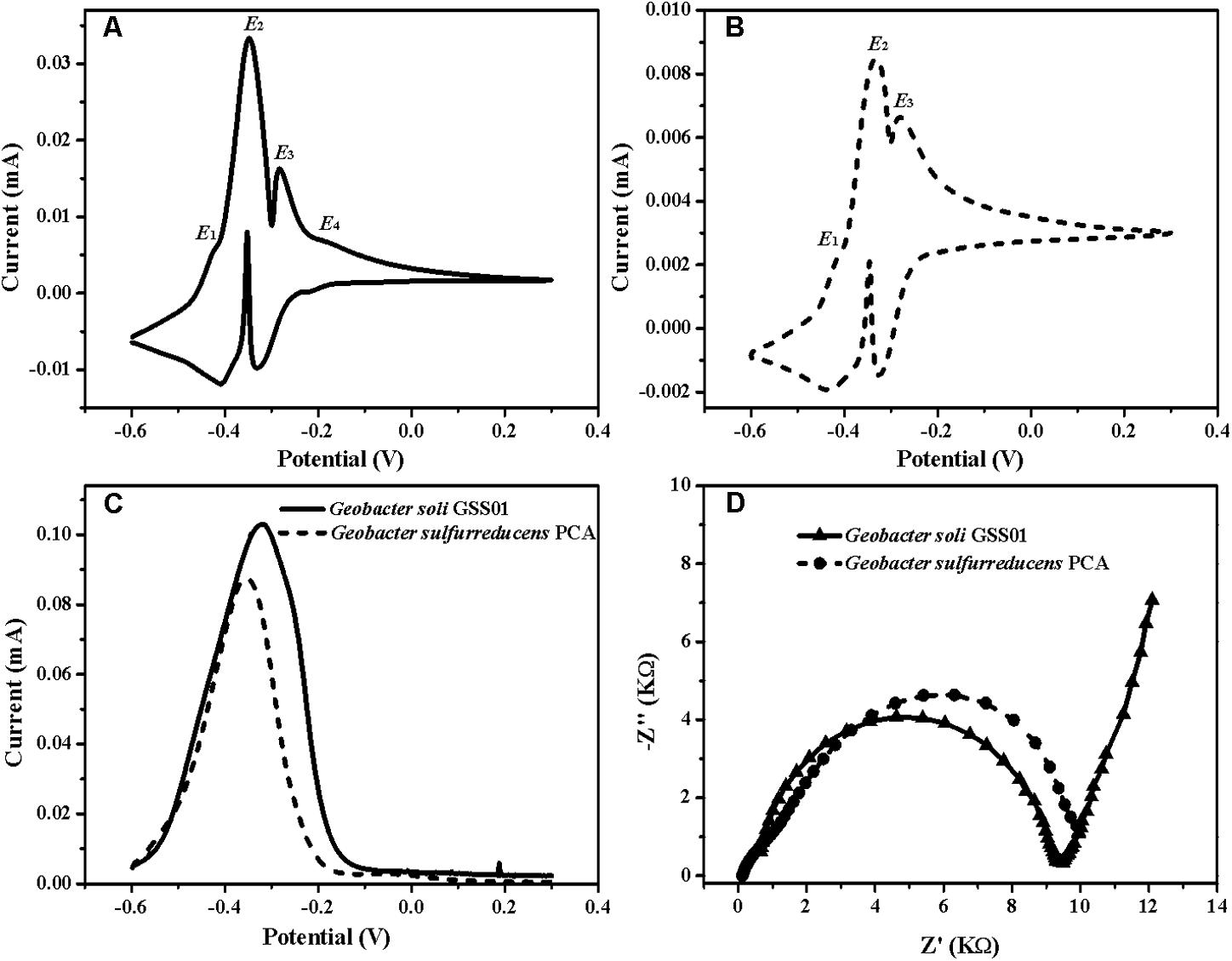
图2 G.soli GSS01和G.sulfurreducens PCA 生物膜电化学结果
3. 外氧化还原蛋白的电化学原位 FTIR光谱
作者采用电化学原位 FTIR光谱获得更多关于G.soli细胞表面蛋白的信息,并在分子水平上监测它们在电化学反应过程中的构象变化。结果显示,对于用FC和ITO生长的G.soli细胞,在1552、1458、1400、1248和1150cm-1处出现正带。与用在1650cm-1处具有明显条带的FC生长的细胞的光谱相比,用ITO生长的G.soli细胞的光谱在1700-1600cm-1处具有一个更宽的条带。此外,图3 C的带肩从1640移动到1670cm-1,电位从-0.7增加到0.3V,并且在-0.1V 处将该带分成1668和1620cm-1处的两个带。不同光谱振动反映了由电子受体引起的G.soli细胞的外部蛋白质的独特模式。酰胺I带和其他带的显著差异表明,在G.soli和G.sulfurreducens之间有独特的外部氧化还原蛋白促进两种Geobacter菌种的电化学行为。由于具有较高氧化还原电位的外表面蛋白将促进电子从周质向外传递,作者推测额外的氧化还原蛋白一方面可能有助于提高G.soli的EET能力,另一方面表明在G.soli中可能存在未知的电子传递导管。
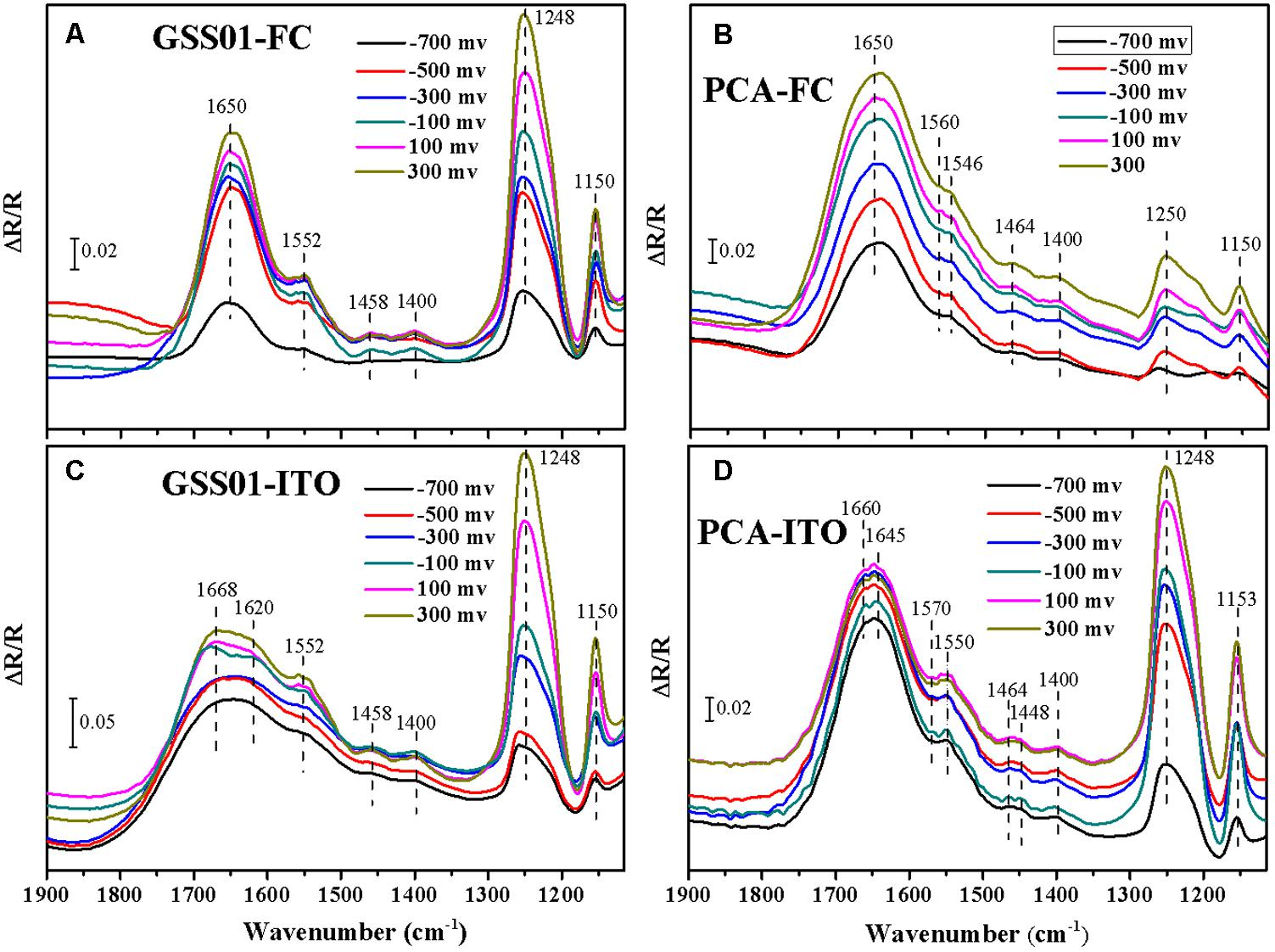
图3 电化学原位 FTIR光谱分析结果
4. 转录组学和蛋白质组学分析
在转录组学结果中,作者分别鉴定到了2013(FH vs FC)、1705(ITO vs FC)差异基因(Pvalue≤0.05),在蛋白质组学结果中,共鉴定到了2149个蛋白质,差异蛋白数分别为409(FH vs FC)、207(ITO vs FC)(FC≥1.5,Pvalue≤0.05)。作者发现,许多在转录组学分析中具有较高转录水平的基因,例如cbcL、imcH、rplE和rplN,在蛋白质组学分析中具有降低的翻译水平,这可能是因为膜相关蛋白的低提取产率或表达的转录后调节。
5. 与代谢和生长相关的蛋白质
在本次研究中,作者选择了与三羧酸(TCA)循环、氧化磷酸化和翻译相关的基因、蛋白质的表达水平进行了分析。
在TCA循环中,使用FH和ITO的细胞生长期间,与代谢率直接相关的柠檬酸合酶(GltA)的表达水平在转录组学和蛋白质组学分析中均下调。在转录组学分析中,三种琥珀酸脱氢酶编码基因(frdA,frdB和frdC)、苹果酸脱氢酶(mdh)和异柠檬酸脱氢酶(icd)编码基因的表达水平下调。TCA循环中许多基因的转录水平,如frdB和乌头酸氢盐编码基因acnA,在用ITO生长的细胞中的蛋白丰度比在用FH生长的细胞蛋白丰度更低,说明用ITO生长的细胞中的代谢率较低。
在使用FH或ITO细胞生长期间,催化呼吸代谢反应酶的调节是复杂的。在蛋白质组学分析中,F0F1型ATP合酶的一个亚基(AtpH)在用FH生长的细胞中比在FC中表达量上调,而另一个亚基(AtpG)则表达量下调;在转录组学分析中,细胞色素c氧化酶(CoxA、CoxC和CoxD)的表达上调,细胞色素bd甲基萘醌氧化酶(CydA)的表达下调。这和已有的G.sulfurreducens研究结果一致,即两种NADH脱氢酶复合物在所有测试条件下均表达,但不同亚基的丰度是不同的。一般来说,大多数亚基在FH生长过程中比FC更丰富,但在ITO生长过程中丰度低于FC,这也表明使用ITO生长的细胞代谢率低于FH。
更多的核糖体产生表明微生物有更快的生长速率。蛋白质组学分析中,许多核糖体蛋白在FH和ITO培养的细胞中的丰度比FC低约1.5-25倍,这支持了不溶性电子受体细胞生长较慢的结论。然而,大多数这些核糖体蛋白编码基因的转录水平在用FH和ITO生长的细胞中比在FC中更丰富。该结果表明核糖体蛋白的表达高度依赖于如前所述的转录后控制。
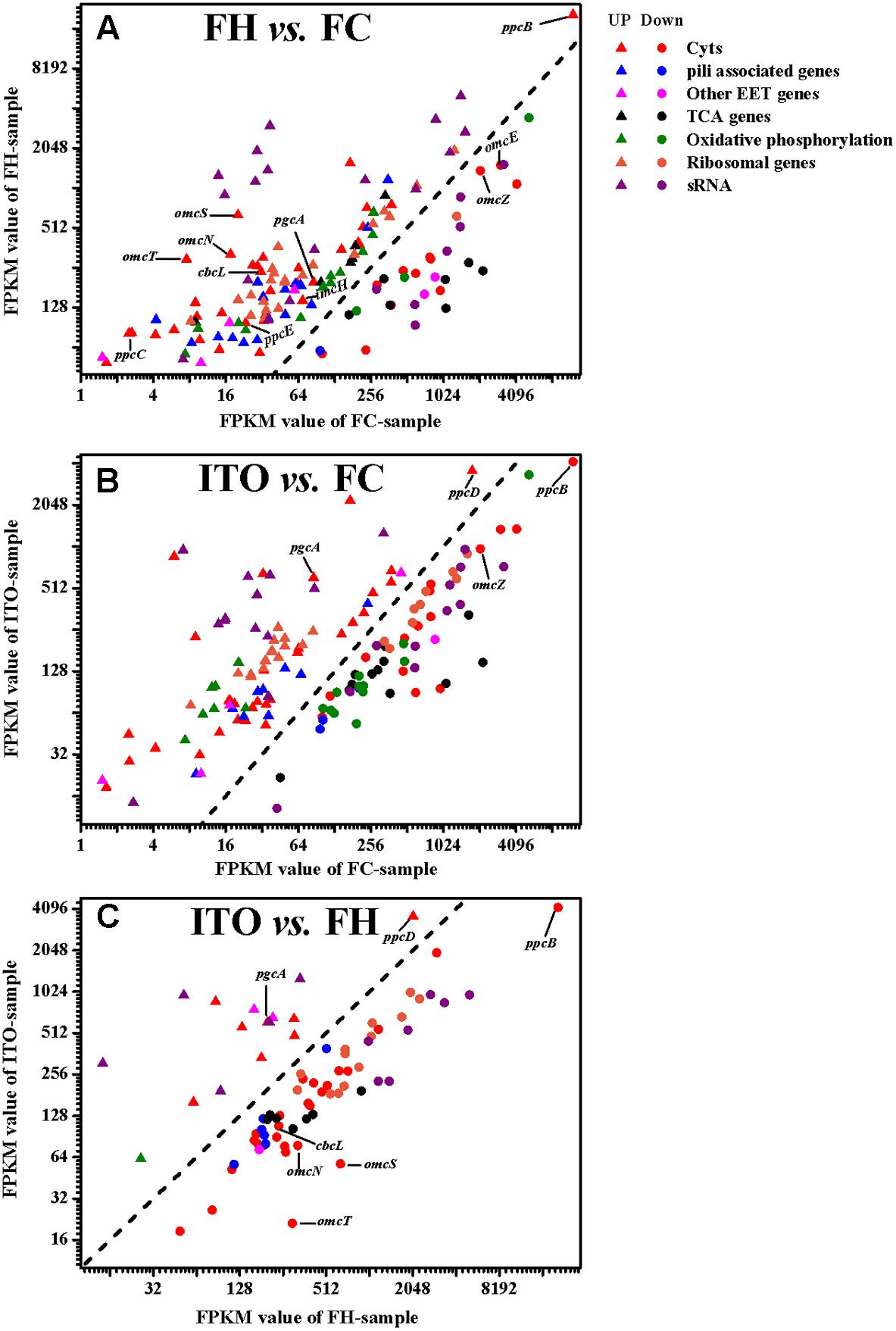
图4 G.soli细胞转录表达水平比较
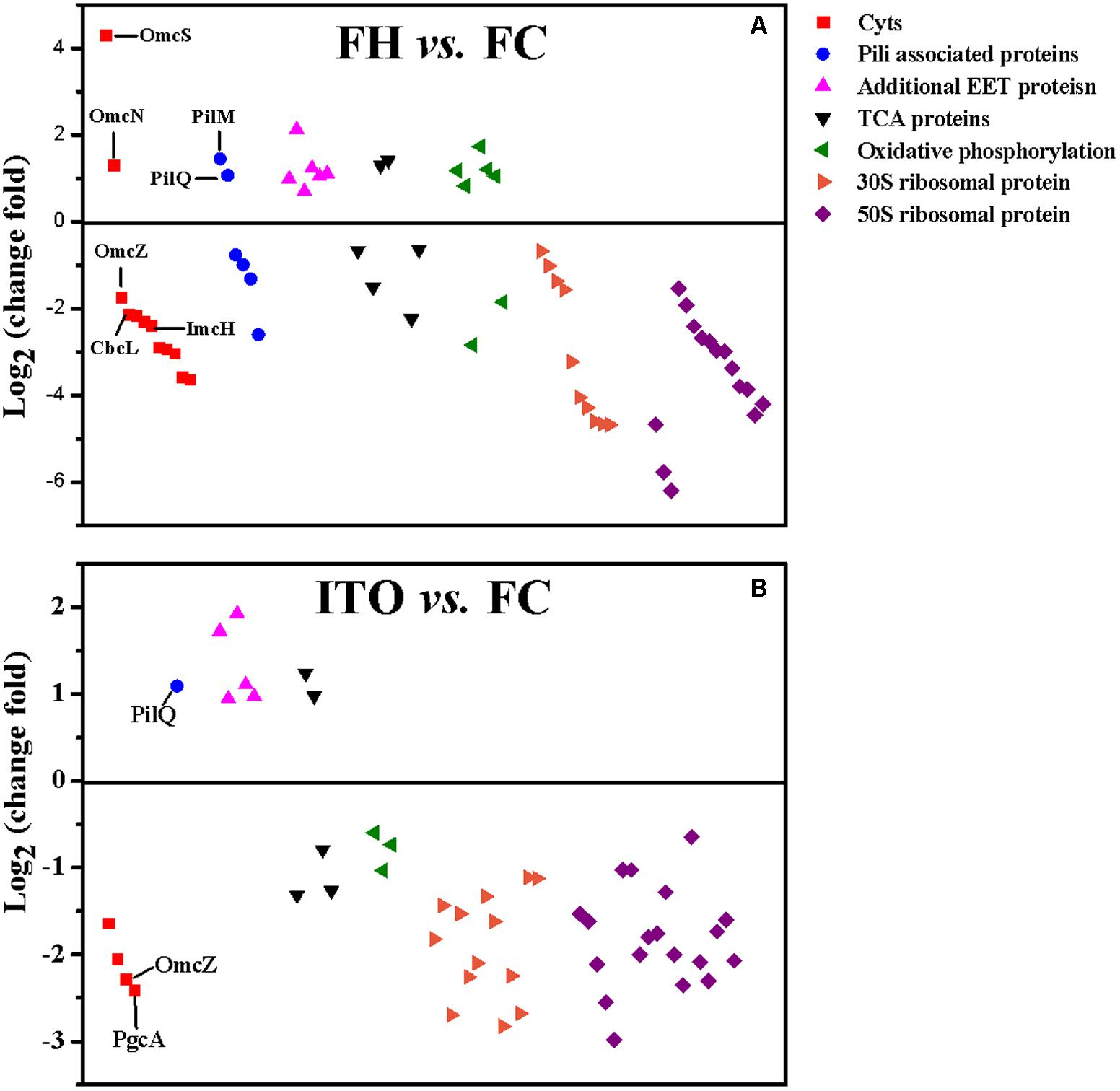
图5 G.soli细胞蛋白表达水平比较
6. 可能参与EET的蛋白质
作者认为可能参与EET的蛋白主要包括细胞色素蛋白、菌毛相关蛋白、额外的电子转移蛋白、小的非编码RNA(sRNA)等。
目前来自G.sulfurreducens研究表明,许多c-Cyt蛋白在EET中起重要作用:(1)内膜电子转移至少通过两种不同的途径,分别为CbcL-依赖途径和ImcH-依赖途径;(2)从内膜到外膜的电子转移可能是由PpcA家族(PpcA-PpcE)介导;(3)通过外膜转移电子依赖于反式外膜孔;(4)从外膜到电子受体的末端电子转移由OmcS和OmcZ介导。
在转录组分析结果中,共有62个c-Cyts显著差异编码基因,其中有23个在两组中表达皆上调。基因cbcL和imcH的表达在用FH生长的细胞中分别上调7.6和2.1倍,而在用ITO生长的细胞中没有明显变化。在蛋白质组学分析中,蛋白质cbcL在用FH生长的细胞中表达下调。由于FC,FH和ITO的氧化还原电位彼此不同,含有FC和FH的培养物的氧化还原电位将随着Fe(III)还原而降低,说明这两种蛋白质的调节不仅直接依赖于电子受体的类型,而且还依赖于培养物中Fe(III)-还原比率。
在五个密切相关的小型三聚体c -Cyts PpcA家族中,ppcB、ppcC和ppcE的转录水平在用FH生长的细胞中上调,而ppcD在用ITO生长的细胞中比在FC中具有更高的转录水平。作者推测这些蛋白质均通过PSORT进行周质定位。
omcB基因在FH和 ITO生长的细胞中的转录水平均上调,但它的翻译水平在蛋白质组分析结果显示并没有明显变化。在蛋白质组学分析中,用FH或ITO生长的细胞中两个表达上调的蛋白的c-Cyts是SE37_02820和SE37_11760,而在G.sulfurreducens中最接近SE37_02820的同源物是OmcS,具有94.7%的氨基酸序列同一性。之前G.sulfurreducens中的研究证实,OmcS是末端Fe(III)氧化物还原酶,但对于电极还原不是必需的。OmcZ在促进从生物膜到电极表面的电子转移方面发挥重要作用,已发表的研究结果认为omcZ的转录水平在不可溶电子受体中表达大幅度上调。然而,在本研究中作者发现,与FC生长的细胞相比,在FH和ITO生长的细胞中omcZ(SE37_04285)转录和翻译水平显着降低。
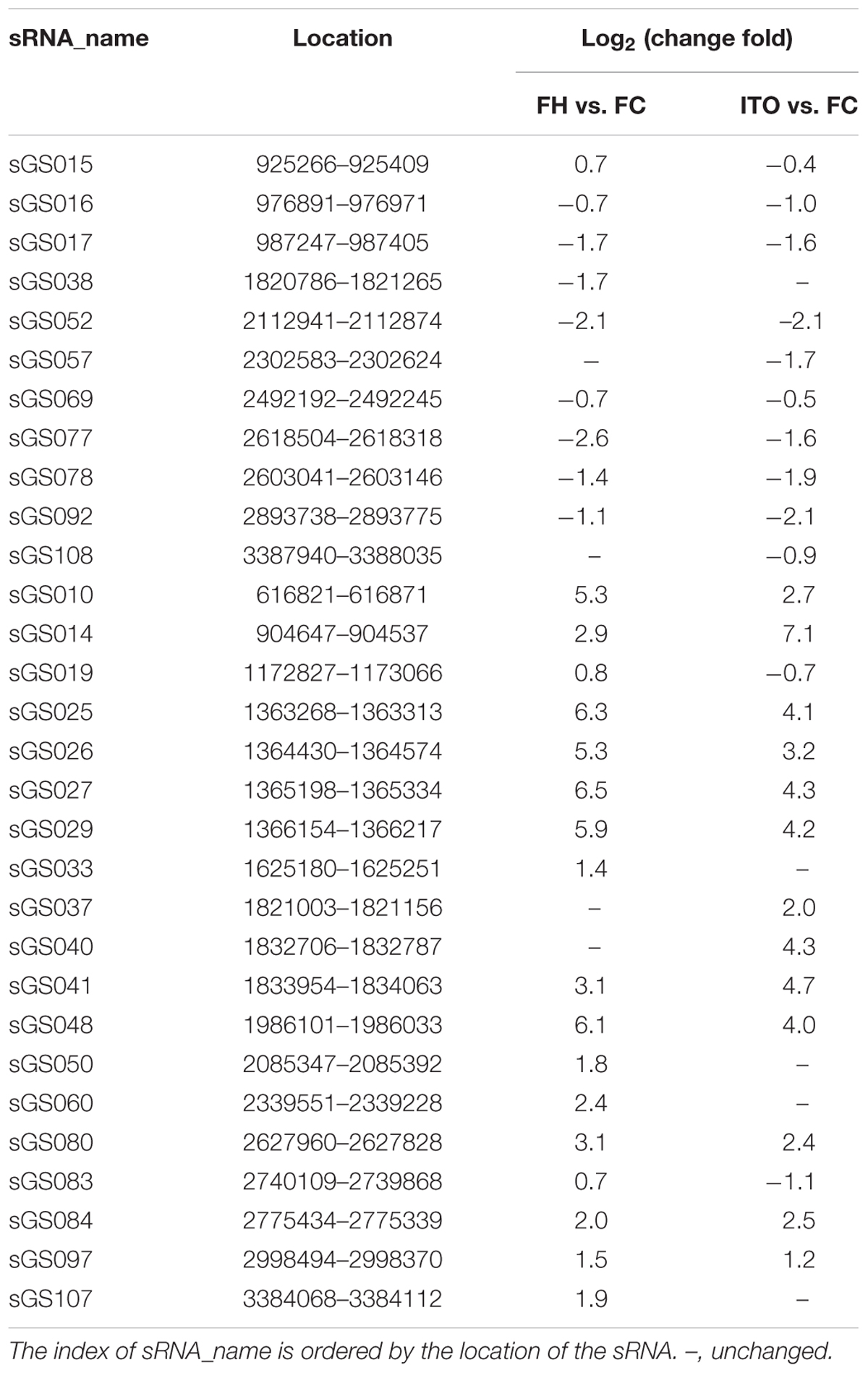
细菌中的许多外膜蛋白都是通过sRNA调节的,而外膜蛋白对EET很重要,因此sRNA的差异表达同样很重要。比较转录组学显示,在用不溶性电子受体和可溶性受体生长的G.soli细胞中差异表达30种新型sRNA,差异表达最大的sRNA是sGS014。为了进一步研究sRNA在EET中的潜在作用,作者使用RNA相互作用预测工具IntaRNA分析差异表达的sRNA和EET基因之间的相互作用,结果显示sGS014可以与30个EET基因相互作用,并且预测两个外膜c -Cyts SE37_11760和SE37_11830与多达28个不同的sRNA相互作用。由于几乎所有鉴定的sRNA都在较低选择压力的基因间区域编码,因此它们在不同的Geobacter物种中具有更大的突变空间。sRNA可参与调节不同Geobacter物种中EET基因的表达,需要进一步研究以确定它们对基因表达的物种特异性调节的影响。
表1部分重要的c- Cyt蛋白和可能参与EET的其他蛋白质
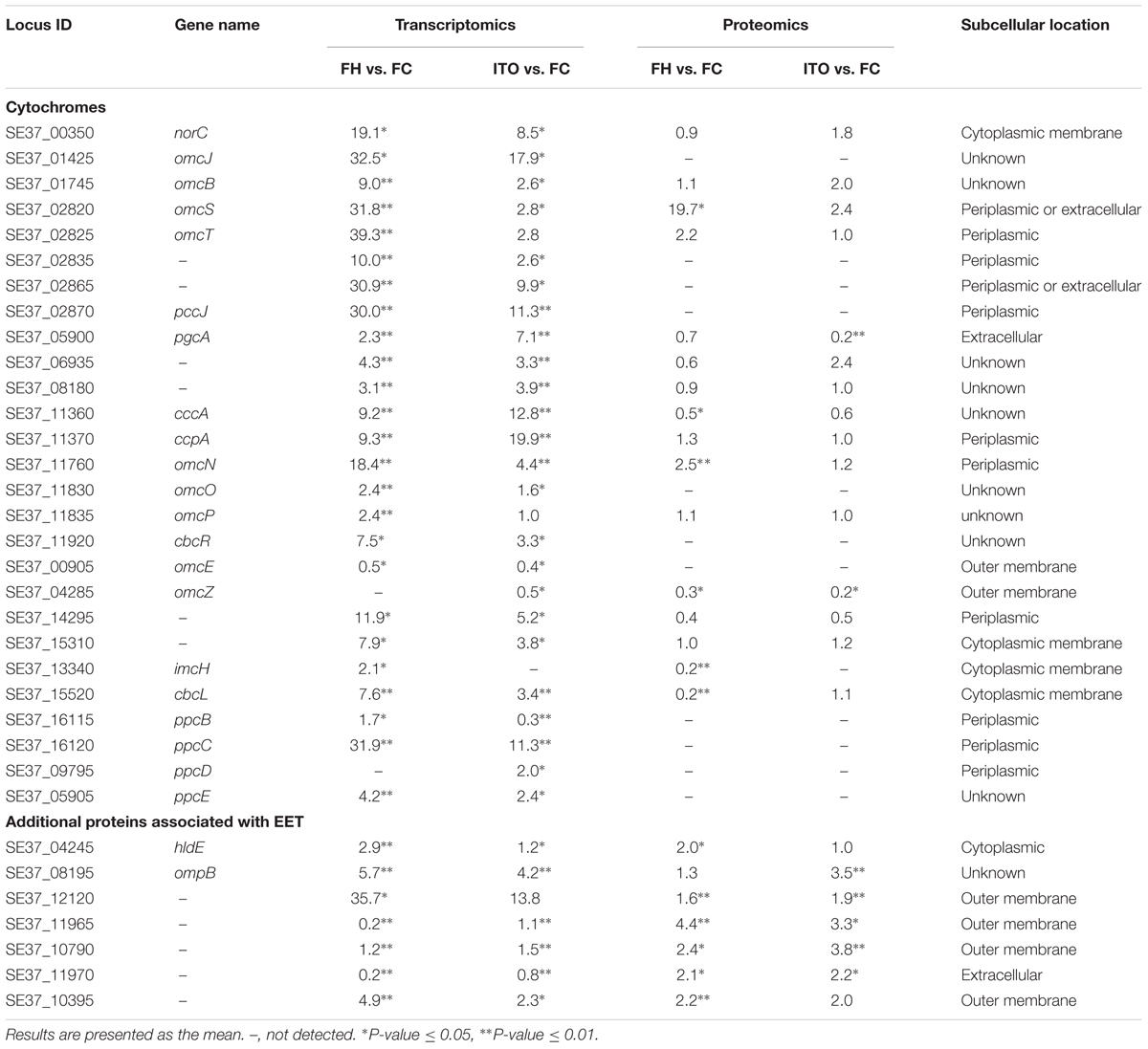
表2 差异表达的sRNA候选物
研究结论
本研究表明,G.soli是一种有潜力的Fe(III)氧化物还原剂和电流产生剂,其EET机理值得进一步研究。在用不溶性电子受体和可溶性电子受体生长的G.soli细胞中鉴定到具有显着差异丰度的蛋白质集,可用于将来对该物种中EET机制的研究。sRNAs在Geobacter属中调节EET基因表达的可能性为探索EET机制提供了新的理论基础。
解析文献
Xixi Cai, Lingyan Huang, et al. Transcriptomic, Proteomic, and Bioelectrochemical Characterization of an Exoelectrogen Geobacter soli Grown With Different Electron Acceptors. Frontiers in Microbiology , 2018 , 9 :1075.
参考文献
1. Ding, Y. R., Hixson, K. K.,et al. (2008). Proteome of Geobacter sulfurreducens grown with Fe(III) oxide or Fe(III) citrate as the electron acceptor. Biochim. Biophys. Acta 1784, 1935–1941.doi: 10.1016/j.bbapap.2008.06.011
2. Yang, G., Chen, S., Zhou, S., and Liu, Y. (2015). Genome sequence of a dissimilatoryFe(III)-reducing bacterium Geobacter soli type strain GSS01T. Stand. GenomicSci. 10:118. doi: 10.1186/s40793-015-0117-7
3. Merkley, E. D., Wrighton, K. C., et al. (2015). Changes in protein expression across laboratory and field experiments in Geobacter bemidjiensis. J. Proteome Res. 14, 1361–1375.doi: 10.1021/pr500983v
4. Kavanagh, P., Botting, C. H., et al. (2016).Comparative proteomics implicates a role for multiple secretion systems in electrode-respiring Geobacter sulfurreducens biofilms. J. Proteome Res. 15, 4135–4145. doi: 10.1021/acs.jproteome.5b01019
下一条:慢病毒包装经典文献解析
最新动态
-
03.11
同为抗体,为什么一抗看“抗原种类”,二抗却看“一抗种属”?
-
03.11
J Nanobiotechnology(IF=12.6):蔓越莓“纳米邮差”递送麦角甾醇,重启早衰卵巢功能!
-
02.27
没有抗体?不是模式植物?DAP-seq照样搞定你的TF靶基因!
-
02.27
高分研究的共同选择:三篇权威文献背后的ChIP实验工具解析
-
02.27
表观遗传学进入“多维验证”时代,这些疾病机制再也藏不住了!
-
02.27
“药食同源”顶刊思路(IF=10.5)!来源鉴定+活性追踪+体内验证,全面解析无花果来源外泌体抑制乳腺癌骨转移的双重机制!
-
02.27
千年古方,纳米新生:科学家破解半夏如何“指挥”免疫细胞对抗肺癌!
-
01.29
技术跨界联用:表观遗传学进入“多维验证”时代,这些疾病机制再也藏不住了!文末有惊喜活动!
-
01.29
机制研究的金标准:金开瑞RIP/RNA pull-down试剂盒获多篇顶刊研究认可!
-
01.29
想测结合常数,但样本太珍贵?来试试MST技术
 X
X 






